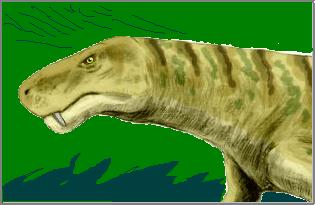
Синапсиды.
Для Вас:
Общая характеристика.
Синапсиды – класс амниот, т.е. наземных четвероногих (хордовых) животных, включающий прямых предков млекопитающих, а также родственные этим предкам формы. Ранее синапсид причисляли к классу рептилий (пресмыкающихся), но синапсиды гораздо дальше отстоят от современных пресмыкающихся, чем от млекопитающих. Возможно, синапсиды и диапсиды (главная ветвь рептилий) происходят от одного предка, которого уже можно отнести к амниотам, но возможно и то, что синапсиды и рептилии независимо произошли от земноводных (находящихся, однако в близком родстве). По крайней мере, достоверно известно, что эволюционные ветви синапсид и рептилий окончательно разошлись в позднем карбоне около 320 млн. лет назад. И эволюция этих ветвей амниот происходила по-разному и даже в разных коиматических условиях.
Начиная с конца каменноугольного периода, синапсиды развивались в сторону интенсификации обмена, активного перемещения и, в конечном итоге, теплокровности. А эволюция рептилий в этот период была направлена в сторону независимости от источников воды и экономии энергии. Видимо, по этой причине в течение перми произошло разделение зон обитания синапсид и рептилий: синапсиды населяли влажные районы, рептилии, в основном, проживали в аридных приэкваториальных и тропических областях.

Хорошо сохранившийся и частично восстановленный череп Диметродона. Отлично заметна разница в размерах зубов в верхней и в нижней челюсти. Хорошо просматриваются швы черепа. Также стоит отратить внимание на относительно небольшой размер височных окон и отсутствие рельефа на их краях.
На картинках слева у нас эволюция пермских синапсид от раннепермского хищного пеликозавра Диметродона до позднепермского хищного тероцефала Ликозуха и до позднепермского травоядного Дицинодона.
Основной классификационный признак синапсид – череп с одним височным окном, предназначенным для крепления приводящих челюстных мышц. Видимо, такая конструкция челюстного аппарата оказалась очень удачной, поэтому среди синапсид появилось большое количество хищников, обладавших мощными и достаточно длинными челюстями, которые позволяли захватывать добычу, удерживать ее и при этом наносить тяжелые увечья. В этом хищным синапсидам помогали также крупные клыки, которые также стали визитной карточкой синапсид. Дифференциация зубов вначале была только по размеру; уже у ранних синапсид выделились крупные клыки и более мелкие передние и заклыковые зубы. Пределом дифференциации по размеру стало полное исчезновение заклыковых зубов у позднепермских горгонопсий. Но примерно в это же время у цинодонтов заклыковые зубы получили жевательную функцию и стали превращаться в коренные, т.е началась дифференциация зубов по форме. Одновременно началось формирование челюстного сустава характерного для млекопитающих. Вполне возможно, что в это же время синапсиды обзавелись и губами, которые в последствии облегчат жевание, а также позволят сосание молока и обработку пищи слюной в ротовой полости. В течение пермского периода синапсиды сохраняли небные зубы, но их количество и функционал сокращались в плоть до полного исчезновения у самых совершенных тероцефалов и цинодонтов.
Развитие челюстных мышц и появление крупных клыков резко увеличило нагрузку на челюсти, и для того чтобы увеличить прочность верхней челюсти, на которую приходится большая нагрузка, у синапсид к концу пермского периода развилось вторичное костное небо. Параллельно вторичное небо позволяло дышать при нахождении пищи в ротовой полости.
Кожа синапсид, скорее всего, была чем-то средним между кожей земноводных и кожей млекопитающих. То есть скорее всего первоначально (у пеликозавров) это была тонкая кожа, насыщенная железами и вполне проницаемая для воды и газов. Скорее всего, кожное дыхание было важной частью газообмена. Также, скорее всего, существовало ороговение внешнего слоя кожи, но если кожа и была покрыта чешуей, то эта чешуя напоминала чешую крокодилов, но была менее крупной и менее развитой. У позднепермских форм, возможно, появляются зачатки волосяного покрова. Хотя пока не понятно, как шло преобразование чешуи в волос, или как появились железы, постоянно вырабатывающие кератин и формирующие волос.
За важность кожного дыхания для синапсид говорит то, что у них до конца пермского периода сохранялся примитивный рептилийный тип дыхания. Диафрагма появилась лишь в триасовом периоде.

Фотография черепа позднепермского тероцефала Ликозуха (Lycosuchus) сверху. Хотя многие детали строения, в частности зубной ряд, на этой фоторгафии не видны, зато заметны отличительные особенности строения заглазничной части черепа тероцефалов: видно, что височные окна очень велики; крышка черепа в задней части редуцирована (остался лишь гребень между правым и левым окном); перемычка между глазницами и височными окнами очень узкая. Интересен и слабо вогнутый затылок...
Ранние синапсиды напоминали по внешнему виду и способу передвижения ящериц, однако в ходе эволюции на протяжении пермского периода конечности оказались заметно подвернуты под туловище, а передвижение стало похожим на движение современных млекопитающих рысью. При этом у даже у поздних пермских синапсид сохранялись мышцы, используемые при передвижении в стиле рептилий. Плечевой пояс синапсид сохранял конструкцию типичную для пресмыкающихся, но кости плечевого пояса изменились и перестроились, чтобы передние конечности были более-менее выпрямленными. С таким устройством плечевого пояса было еще очень далеко до передвижения галопом. Более того, при движении шагом или рысью животные, во-первых, опирались на всю кисть; во-вторых, они сильно изгибались и раскачивались из стороны в сторону; в-третьих, их походка была косолапой. Однако нужно заметить, что частичное выпрямление конечностей было характерным только для некрупных форм. Более того, в течение пермского периода, хотя задние конечности синапсид приобрели частично выпрямленную постановку (за счет изгиба), у синапсид не модифицировался тазовый пояс и не изменилась головка бедренной кости, чтобы синапсиды смогли выпрямить задние конечности, как это сделали архозавры уже в середине триаса. Более того, синапсиды, в отличие от триасовых архозавров, сохранили примитивное устройство стопы (с двойным голеностопным суставом и без выраженной пятки), которая была только опорой, не служила для удлинения конечности или усиления толчка при беге. Однако к концу пермского периода стопа у синапсид ставилась параллельно туловищу, и самым длинным стал 3-й палец.
Первоначально синапсиды не имели ни наружного, ни среднего уха. Внутреннее ухо и органы равновесия они унаследовали от амфибийных предков, но по мнению многих специалистов, это внутреннее ухо могло лишь улавливать вибрацию почвы, если животное ложило на землю нижнюю челюсть. Но, как считается, в средней перми задняя часть нижней челюсти стала выполнять роль резонатора воздушных колебаний, в конце перми, пожалуй, появилась барабанная перепонка, однако передача звука шла через челюстной сустав, что было неэффективно.
Эволюция синапсид в пермском периоде.
Заметим, что эволюция синапсид даже в период их господства – в первую очередь эволюция неспециализированных хищников и всеядных. И уже от них ответвлялись другие эволюционные направления.
Основное направление в эволюции синапсид связано с развитием челюстного аппарата. Сначала раскрыть рот как можно шире, затем добиться максимальной силы укуса (за счет появления новых мышц и клыков), после научится перетирать (как дицинодонты) пищу или жевать ее (как цинодонты). Все это означало уменьшение длины морды относительно заглазничной части черепа, но одновременно увеличение ее ширины, высоты и прочности.
В ходе эволюции синапсид к концу перми возникло три типа челюстного сустава, каждый из которых своеобразно решал указанные выше задачи. Имеются в виду челюстной сустав горгонопсий с подвижным квадратом, дающий максимально широкое раскрытие рта; скользящий сустав у дицинодонтов, гарантирующий наилучшее перетирание пищи; переходный к маммальному челюстной сустав цинодонтов, рассчитанный на жевание.
В эволюции синапсид подвижность играла наименьшее значение. Хотя постепенно шло выпрямление конечностей, но даже у самых совершенных синапсид в конце перми сохранялось типичное для пресмыкающихся расположение конечностей, а также примитивные запястные и голеностопные суставы. Более того, в течение перми не раз более подвижние синапсиды проигрывали менее подвижным, но обладающим более совершенными челюстями.
Зато движение к теплокровности было одним из основ трендов эволюции синапсид. Дело в том, что они активно осваивали умеренный климатический пояс, в котором жить приходится при сильных сезонных колебаниях температур. А в этом случае теплокровный хищник всегда выиграт у малоподвижных холоднокровных жертв.
Группы синапсид.
В ранней перми преобладали более примитивные пеликозавры, появившиеся еще в Карбоне, в средней и поздней перми сменилось несколько генераций синапсид, относимых к более прогрессивным формам, имя которым терапсиды. К последней из этих генераций относились цинодонты, от которых в триасе или юре произошли млекопитающие.
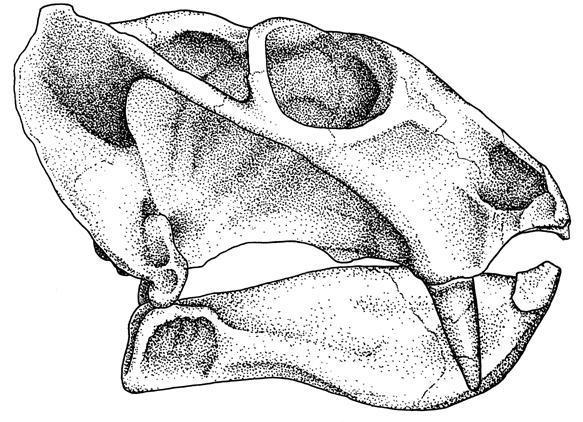
Рисунок, изображающий череп одного из представителей рода Dicynodon, относящегося к поздней перми. В первую очередь стоит обратить внимание на строение заглазничной области черепа с ярко выраженными височными арками, которое уникально для Синапсид. Также стоит обратить внимание и на челюстной сустав. А именно у этого образца желательно взглянуть на низкорасположенные ноздри, из-за которых животное вряд ли могло дышать в процессе перетирания пищи.
Поскольку синапсиды были доминирующей группой позвоночных в пермском периоде, то количество видов, родов, семейств и других систематических групп синапсид перми просто огромно. Обычно, рассказывая о них, рисуют дерево кладограммы, показывающей генетические взаимосвязи групп животных. Но мы пойдем другим путем: укажем экологическую нишу и посмотрим какие животные занимали ее в разные эпохи пермского периода. Итак, нишу доминирующих хищников занимали:
- В ранней перми -- сфенакодонты и офиакодонты, первые были чисто сухопутными хищниками, а вторые – (вероятно) полуводными.
- В начале Гваделупской эпохи в восточной части субэкваториальной зоны доминирующими хищниками были биармозухии (и родственные им эотитанозухии)
- Всего через пару миллионов лет на смену биармозухиям пришли Антеозавры -- хищные диноцефалы.
- В поздней перми, после вымирания диноцефаловой фауны роль главных наземных хищников досталась Горгонопсиям, ставшим последней доминирующей палеозойской группой хищных синапсид.
А теперь мы перейдем к крупнейшим травоядным синапсидам перми:
- В начеле Приуральской эпохи основными травоядными были эдафозавры, известные из поднего карбона и ранней перми Евроамерики.
- Но очень быстро эдафозавры были сменены касеидами (иначе Казеиды, Caseidae).
- Еще во время существования касеид в приуралье (Очерская фауна) распространились эстемнозухи, предположительно первые диноцефалы.
- Диноцефалы продолжали оставаться крупнейшими травоядными по оканчания средней перми. Они были преставлены двумя группами тапиноцефалами и титанозухиями.
- В начале позднепермской эпохи синапсиды ненадолго уступили нишу доминирующих травоядных парейазаврам, но ближе к концу эпохи основными травоядными стали Дицинодонты, не только просуществовавшие до конца перми, но и пережившие Пермско-триасовое вымирание.
Поскольку останки крупных животных хорошо сохраняются, то мы легко можем сказать, какие виды были основными хищниками и основными травоядными в том или ином сообществе. Гораздо труднее определить какие животные были промежуточными звеньями пищевой цепи. Дело в том, что, например, насекомоядные и мелкие хищники, как правило, сами становились добычей более крупных хищников и падальщиков. К тому же мелкие специализированные хищники и специализированные насекомоядные вынуждены были конкурировать с недостигшими зрелости особями крупных хищников. По крайней мере, можно перечислить следующие группы синапсид, являвшиеся промежуточными хищниками и насекомоядными:
- В течение ранней и (возможно) средней перми успешной группой мелких хищников были Варанопсеиды (Varanopseidae). (В некоторых природных комплексах варанопсеиды могли быть и доминирующими хищниками.)
- В средней перми известны следующие некрупные хищники: Stenocybus (роудский век); бурнетиаморфы (Burnetiamorpha) жившие в конце средней - начале поздней перми; а также иктидорхиниды (Ictidorhinidae) известные из начала поздней перми, но очевидно существовавшие и ранее.
- В поздней перми эту нишу заняли две крупные группы животных Тероцефалы (Therocephalia) и Цинодонты (Cynodontia).
Кроме мелких хищников из пермского периода известны и мелкие травоядные. Однако, известно их немного, и, скорее всего, синапсиды в пермском периоде были неготовы занять эту нишу. В средней и поздней Перми в обоих полушариях существовали аномодонты (Anomodontia), считающиеся родственниками и предками дицинодонтов.

Все, что нужно, можно найти прямо здесь:
|
|
-- --
Парк Пермского Периода 2010-2022